蜘蛛全身分布着高度敏感的机械感受器,能够分辨微风和猎物挣扎
图文|桦语侃史
编辑|桦语侃史
前言:
蜘蛛的机械感知器官,这是最敏感的机械感受器之一。通过了解蜘蛛机械感知器的分布、形状和功能,以及竖琴状感知器的结构。然后结合机械感知器的神经元电生理特性,可以很好的解决

从单细胞的生物体到大多数脊椎动物,机械感觉都是一项非常重要的功能。它是触觉、平衡感觉、听觉和肌肉运动感知的神经调节基础。
它为运动控制和自稳态的维持提供重要的反馈信号,如血压和呼吸节律的调节。蜘蛛被描述为丑陋的生物,具有8条腿和吐丝结网的能力。大部分时间蜘蛛躲藏在蛛网后面,夜间则呆在蛛网中央守株待兔,等待猎物自投罗网。目前已发现的蜘蛛种类达到3.5万多种,分布广泛,几乎在全球各地都可以看到蜘蛛。
蜘蛛以吐丝结网和用毒液捕食而闻名。令人惊叹的是蜘蛛全身分布着高度敏感的机械感受器,蜘蛛的机械感受器是最灵敏的之一,能够分辨微风和猎物挣扎之间微小的差异!20世纪60年代,美国海军研制出了一种名为“海底蜘蛛”的机器人,用于密切监视水下情况。

蜘蛛的机械感受器在仿生学上有着巨大的应用价值。如今,研究人员对蜘蛛在仿生学上的应用越来越感兴趣,根据蜘蛛绒毛的敏感性和精确性,Barth和Holler设计了纳米生物传感器。
此外,Albert正在研制一种类似于蜘蛛的隐性网络雷达系统,以便能够像蜘蛛一样根据蛛丝运动的变化来采取行动。近年来,研究人员主要使用免疫组织化学、电子显微镜技术和电生理技术来揭示蜘蛛机械感受器的结构和功能。
蜘蛛具有高度进化的感受器,这使它能够在自然环境中良好地生存和发展。Jiang等人使用扫描电子显微镜观察到,蜘蛛的机械感受器分布于全身。头部覆盖着三明治状和半球状的压力感受器,腹部的压力感受器呈现三明治状。

三明治状的压力感受器具有较高的敏感性,而半球状的主要起缓冲和保护作用。压力感受器上长有几微米长的绒毛,尖部的直径是中部的1/20,约为200nm,尖部的敏感性很高,中部具有较高的柔韧性和阻抗性。
绒毛主要分布在腿部,腿上的绒毛上生长着两种类型的刚毛。一种是短而圆柱状分布在附骨的远端,用于感知较大的气流;另一种是纺锤形的,其质量只有10g。实验证明,该种刚毛引发神经冲动所需的最小机械能为4x10-2Ws,比最敏感的光感受器还要灵敏100倍。
这表明其敏感性有多高,任何微小的风吹草动都能被它感知到。每条腿上通常有1对爪,2爪之间的距离为10微米,爪上的枝状指间的距离为500nm至2um,正好可以使1至4根蛛丝通过,蛛丝的振动使爪能够感知。
蜘蛛表皮上分布着竖琴状感受器,对张力敏感,用于感知蜘蛛自身运动产生的张力和压力,以及猎物、天敌和配偶所发出的震动。各种机械感受器之间相互协调、密切配合,通过感受器感知周围环境的变化,将信号传送到中枢,由神经中枢进行综合,得出判断,然后做出相应的反应)。
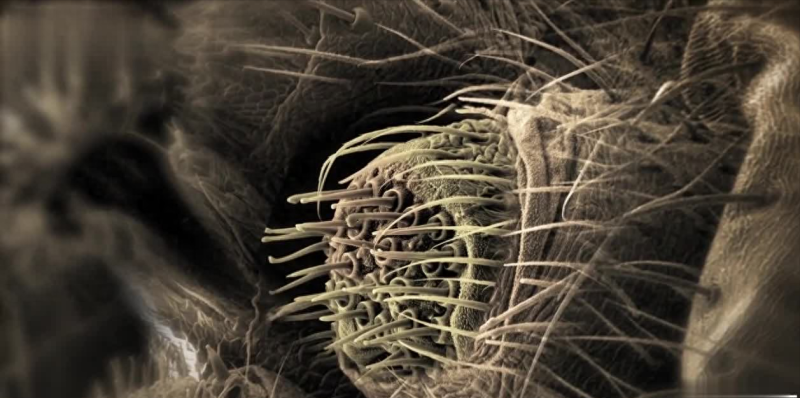
竖琴传感器的结构
近年来,人们对位于腿部膝盖处的竖琴状感器进行了大量的研究,因为这个部位具有较大的感觉神经元。这些感觉神经元的胞体距离感觉转换部位较近,在进行感觉转换同时还能够进行细胞内记录。研究发现,竖琴状感器由7~8条并行排列的沟组成,看起来就像竖着放的琴。
通过超薄切片和电子显微镜的观察,发现每条沟的下方都连接着一对神经细胞,这种神经细胞被称为双极神经元。这两个神经元相邻紧密,它们的胞体、轴突和树突都被神经胶质细胞包裹着。胞体的长度超过100m,而树突的长度可达270pm。两个神经元的树突长度不同,其中长树突终止在表皮的外膜,而短树突终止在内膜。树突上覆盖着许多微管。
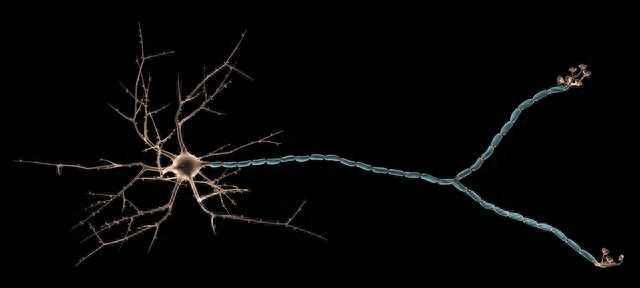
经过实验发现,消除微管后神经元的动作电位和静息电位都有所降低,但动作电位的时程并没有发生改变。因此,微管对动作电位的产生几乎没有影响。目前,微管在竖琴状传感器中所起的作用还不清楚,需要进一步的研究。
感器神经元的电生理特性
细胞内记录表明,支配每条沟的2个神经元动作电位的产生模式不同,被分别称为A型神经元和B型神经元。A型神经元是快适应神经元,轻微的刺激就会产生动作电位,但长时间的刺激仍然只会产生1~2个动作电位。B型神经元是慢适应神经元,但长时间的刺激会使其产生一连串的动作电位。
对鳌虾的实验显示,快适应神经元在牵拉过程中,钙经转导通道内流,激活附近的钙依赖的钾通道,使细胞超极化。蜘蛛的快适应机制可能与此类似。这两种神经元的神经递质也不同,两种神经元都显示乙酰胆碱转移酶免疫阳性反应,因此可能以乙酰胆碱作为神经递质,但只有A型神经元显示强烈的组胺和乙酰胆碱脂酶免疫反应。
即使将感觉神经元的树突和其辅助物质去除,其放电模式也不会改变。因此,感觉神经元的放电模式不依赖于树突和其辅助物质,而是受胞体内部物质的影响。
机械感觉转导的位点
机械感觉通常可以分为三个阶段。首先,机械刺激会作用于感觉细胞的细胞膜,这种刺激会转化为跨膜的电流。这跨膜电流会引发一系列动作电位。研究发现,刺激会作用在感觉神经元树突的不同部位,微电极记录显示,刺激对静止电位没有影响,这意味着机械一电信号转换的部位应该位于树突的顶端。
一般认为,机械转换是通过存在于细胞膜上的压力敏感的离子通道来实现的。在许多组织中,都能发现这种压力敏感的离子通道。然而,尚未直接观察到这种通道在这个过程中的作用。
动作电位产生的起始位点
传统上认为动作电位的产生是在神经元的胞体内进行的。然而,昆虫表皮上的机械感受器的实验显示,树突也具有兴奋的能力。

免疫组织化学研究发现,在整个感觉神经元中都存在着电压依赖的钠离子通道,树突和轴突上的通道密度相同,是胞体密度的两倍。此外,从刺激开始到产生动作电位所需的时间,机械刺激比电刺激要少0.5毫秒以上。这暗示动作电位产生的起始部位靠近树突末梢的感觉转换区域。
受体通道的性质
使用电压钳和河豚毒素的实验发现,细胞上的受体通道对钠离子具有很高的选择性。通过21X射线微量分析和离子敏感电极记录,发现细胞间隙中钠离子的浓度至少和细胞外液中的一样高。
这与许多昆虫感受器细胞和脊椎动物的毛细胞不同,它们具有高浓度的钾细胞外液。当用胆碱代替细胞外液中的钠离子时,电流的产生被阻止。实验还发现,除了钾离子之外,所有其他阳离子都不能通过,仅有50%的钾离子可以通过。
抑制钠离子通道也可以抑制电压。受体通道对pH值也很敏感,在酸性环境下容易开放[26]。受体通道对温度也敏感,与一些脊椎动物和无脊椎动物相似。然而,对于受体对温度敏感的原因和作用机制目前还不清楚。
通过测量变化的电流,可以确定单个通道的电流以及单个神经元上受体通道的数量。实验结果表明,单个通道的电流为7-7.5 Ps,每个神经元上大约有250-500个离子通道[28]。
输出神经对感觉神经元的调控
单克隆抗体显示,感觉神经元周围存在大量的突触连接,这些连接以点状物的形式沿神经纤维分布,并与感觉神经元的树突和轴突接触。显微分析和三维重构技术发现,这些纤维与感觉神经元和胶质细胞之间存在化学突触连接。
当腿部的神经纤维被切断后的4小时内,大部分被切断的神经纤维都被降解掉,这表明这些纤维是来自中枢神经系统的输出纤维。大多数输出神经纤维显示出GABA(γ-氨基丁酸)的免疫阳性反应。
最近的研究发现,GABA并不抑制树突和胞体处的动作电位,而是抑制轴突处的动作电位。免疫细胞化学显示,GABA离子型受体主要分布在感觉神经元轴突及其接近胞体的部位,因此这些输出神经纤维对感觉神经元产生抑制作用。
此外,还有一些输出神经纤维显示谷氨酸的免疫阳性反应。谷氨酸在脊椎动物中是主要的兴奋性神经递质,在这里却具有抑制作用,但其抑制作用没有GABA强烈。因此,谷氨酸可能扮演较低级的调节作用。
使用章鱼胺抗体显示,感觉神经元上存在章鱼胺受体。当章鱼胺作用于感觉神经元时,动作电位的频率明显增加,而这种效应可被特异性抑制剂普鲁卡因阻断。

因此,在这里,章鱼胺是一种兴奋性的神经调节递质,但尚不清楚章鱼胺是否通过突触连接起作用,因为尚未发现明显标记的章鱼胺。
结语
目前虽然在蜘蛛的机械感器研究方面取得了巨大的进展,但仍有许多未解决的问题。首先是机械感觉转换的具体部位及其分子基础,我们仍然不清楚离子通道的结构是什么样的。其次,我们还没有完全阐明感觉神经元周围输出神经纤维的作用。要解决这些问题,需要提高实验手段。
目前,在仿生学上对蜘蛛的研究主要集中在蛛丝的生产和应用方面,取得了很大的进展。然而,关于将蜘蛛的机械感器转化为仿生应用的研究成果还不多。因此,加快在仿生机械领域中将蜘蛛的机械感器应用于生产的进程是非常必要的。

大家都在看
-

这个冬天去哪玩?沈阳16条冬季游特色线路来啦! 玩雪、滑冰、看雪景美食、洗浴、赶大集……这个冬天在沈阳怎么玩你安排了吗?“冬日雪暖阳 ‘圈’出好风光”沈阳都市圈携手推出100条冬季特色旅游线路承包你的冬日快乐!沈阳市、鞍山市、抚顺市、本溪市、阜新市、辽 ... 机械之最12-20
-

财经聚焦·对话企业掌门人丨一根耐寒电缆的创新突围——对话欧耐特线缆董事长杨振涛 近日,欧耐特线缆集团有限公司自主研发的零下40℃耐寒特种电缆,成功中标某大型项目。位于青海西宁市的生产车间内,董事长杨振涛正带领生产团队敲定年后订单的排产细节。从销售代理公司成长为集研发、生产、销售于一 ... 机械之最12-20
-

新华鲜报丨驻华使节吉林行:点赞“冷资源”里的热活力! 新华社长春12月19日电 题:驻华使节吉林行:点赞“冷资源”里的热活力!新华社记者袁睿、姜明明白山松水裹银装,创新发展腾热浪。12月16日至18日,应外交部邀请,23位驻华大使、外交官及国际组织负责人走进吉林省长 ... 机械之最12-20
-

原机械工业部直属5所全国重点大学,如今怎么样了? 中华人民共和国机械工业部,简称:机械工业部、机工部。它是1952年开始组建的,前后共计分有7个部。1998年并入信息产业部, 之后又并入 工业和信息化部 。在它最辉煌的那些年中,直属高校曾达到了23所,其中不乏如今 ... 机械之最12-20
-

我为什么记住了冬至?“家里还有你阿嫲” 原标题:冬至在南方,冬天的到来总是显得珍稀。冬至有一年最长的夜,不单被称为冬节,地位还堪比过年。小时候老人会说“吃了冬节丸就大一岁”,让我以为冬至这一天的汤圆有什么加速时间的魔力,但其实不过是普通的糯 ... 机械之最12-20
-

智慧的巅峰:揭秘三国时期最神奇的机械发明——木牛流马 在中国历史的长河中,三国时期是一段充满英雄豪杰、谋略智慧的时代。其中,蜀汉丞相诸葛亮以其卓越的政治才能、军事谋略和忠诚精神,成为后世敬仰的楷模。而在诸葛亮众多的发明和谋略中,“木牛流马”无疑是最具代表 ... 机械之最12-20
-
仅一根手指一根脚趾能动,他却和母亲建起一座农场! 如果一个人全身瘫痪,卧病在床,靠呼吸机维持生命,仅有一根手指和一根脚趾能动,他还能做什么?在重庆两江新区木耳镇,进行性肌营养不良患者黎夏通过自学物理、化学、计算机编程、机械、农学、医学等知识,和母亲一 ... 机械之最12-19
-

新华网科技观察丨6G与AI融合会带来什么? 新华网北京12月18日电 题:6G与AI融合会带来什么?新华网记者凌纪伟6G与AI,并非两条并行的轨道。AI赋能6G创新,6G又将AI的触角延伸到各领域。两者融合、相互赋能,构筑起智能时代的数字底座。“十五五”规划建议提 ... 机械之最12-19
-

为世界荒漠化治理提供“中国范本”——探寻中国四大沙漠戴上“绿围脖”背后的故事 新华社北京12月18日电 《参考消息》近日刊发文章《为世界荒漠化治理提供“中国范本”——探寻中国四大沙漠戴上“绿围脖”背后的故事》。全文如下:这条人工生态屏障不仅有效遏制了沙尘南下东进的通道,也为世界干旱 ... 机械之最12-19
-

四川哪里的金子最多? 四川日报全媒体记者 王若晔又又又找到金子啦!12月18日,山东烟台消息,莱州市三山岛北部海域新发现国内唯一、亚洲最大的海底巨型金矿。不仅在山东,连月来,全国多地接连传来探“金金金金金”捷报:辽宁探明全国首 ... 机械之最12-19
相关文章
- 四川哪里的金子最多?
- 十几颗下肚,女子痛到直不起腰!医生查完惊呆:实在太大了!千万别这样吃!
- 钱塘江丨布的突围
- 无人机群飞行规划员、智慧仓运维员……科技催生令人心动新职业
- 中外交流丨镜头下的沙海新绿——从图片展上的照片看新疆带给世界的治沙灵感
- 中国玩具如何“玩转”全球大市场?
- 专科生逆袭!2026机械专业必考8大黄金证书,好就业薪资高!
- 高考志愿填报常识34:中国机械“五虎四小龙”
- 在寒风中飘落的树叶是麻烦还是资源?每年520万吨枯枝落叶去哪了
- 跃升48位!太重再次荣登“中国机械500强”榜单
- Anthropic重磅新研究:当AI采访了1250人,它看见了人类的“职业软肋”
- 废墟上,他们把日子重新拧上弦
- 世界五大军事家第5名:成吉思汗 —— 冷兵器时代最恐怖的战争机器
- “十四五”期间 太原市强化企业创新主体地位 激发创新活力
- 外骨骼机器人“出圈” 行业痛点待解
- 理科专业解读一:从学业到就业,一文搞懂机械类专业!
- 大专生逆袭!2026机械设计与制造专业必考8大证书
- 一级军士长的带兵“三字诀”
- 卖“陪伴”成了生意经?为什么大家都不想独处了
- 四大维度,深度解析2025年中国机械工业500强
热门阅读
-
 1
天下第一暗器暴雨梨花针,传说中的唐门暗器做出来了 07-13
1
天下第一暗器暴雨梨花针,传说中的唐门暗器做出来了 07-13 -
 2
2
-
 3
世界十大大型船舶排名,第一能承重六十万吨! 07-13
3
世界十大大型船舶排名,第一能承重六十万吨! 07-13 -
 4
4
-
 5
5
-
 6
6
-
 7
7
-
 8
8